(Mollusca: Gastropoda: Cypraeidae)
with the description of P. chiapponii n. sp.
Caution: this is an old, largely obsolete article. The book Lorenz, F. 2014. "Monograph of the genus Pustularia (Gastropoda: Cypraeidae)". ConchBooks, Harxheim. 130 pp., 30 plts gives a more up-to-date view on this fascinating genus.
Abstract
The genus Pustularia represents a distinctive unit in
the Cypraeid subfamily Erosariinae. In its species, it is difficult
at first sight to determine anterior and posterior end as a result
of a tertiary bilateral symmetry. The species are very similar
to each other and their taxonomy has been subject to recent debates.
This study clarifies some of the points in question and gives
a comprehensive anaylsis of conchological features on the basis
of populations from selected localities. A new species from the
Philippines is described as P. chiapponii.
Contens
0. Prefatory remark on the shell-shape of the family Cypraeidae
1. General notes on the natural history of the
genus
2. Comments on the taxonomy of
some living Pustularia
3. The conchological
features of Pustularia
4. A new species of Pustularia
from the Philippines
5. The living species of Pustularia
6. The subadult
shells of Pustularia
7. Relation between
locality and average shell-length
8. Colour plates
Acknowledgements
Literature
Prefatory remark on the shell-shape of the family Cypraeidae
An ancient systematic criterum for subdividing the animal kingdom
is the body symmetry. The Radiata comprise radially symmetrical
animals (e.g. Cnidaria). The Bilateria are the large group of
bilaterially symmetrical animals, to which the Tribus Mollusca
belongs. Its orders comprise animals with a longitudinal symmetry-axis
dividing the animal in two almost equally built portions. In
most families of the Order Gastropoda, this symmetry is lost
due to the torsion of the visceral sac and the subsequent coiling
of the shell. These animals have secondarily attained an asymmetrical
appearance, often emphasized optically by special shell-processes
on only one side (e.g. Strombidae).
There are a few Gastropod families in which dorsally one side
of the shell is the reflected image of the other - only basally
the differences in the compositon of the two sides is apparent.
In these cases we have a secondary optical relevation of the
body symmetry, from an asymmetrical to a symmetrical shell.
This phenomenon can be observed in the family Ovulidae, Triviidae
and Cypraeidae, in which the species display a certain degree
of bilateral symmetry in shell-shape and -ornamentation. Good
examples are the genera Luria (e.g. pulchra (Gray,
1824)), Bistolida (e.g. stolida (Linné,
1758)) and, last not least, Pustularia (e.g. bistrinotata).
May be, the optical bilatery escapes the food-scheme of molluscivore
crustaceans which detect food by sight. In their habitat of narrow
coral crevices, the Pustularia-species are usually inaccessible
for molluscivore fish, which search for food by chemical rather
than optical stimuli.
1. General notes on
the natural history of the genus Pustularia
The species of Pustularia are highly specialized, mostly
sublitoral reef dwellers. The peculiar shape of the shell in
all species of the genus make it a well defined systematic subunit
within the family Cypraeidae. The genus Pustularia has
ancestral forms mainly in the Caribbean Oligocene. Comparing
living and fossil species, Pustularia might have evolved
from Proadusta -like forms (fig. 1), which hypothesis
makes the sole survivour of the old genus Proadusta surinamensis
(Perry, 1811) the closest living relative in the Atlantic. In
the Indo-Pacific there is the widespread genus Staphylaea
whose granulose surface and dentition are similar to those of
Pustularia. The relationship of the five living species
of Staphylaea to Pustularia is uncertain, but probably
close. Both genera seem to have evolved from a common pustulose
ancestor.
Living species of the genus Pustularia have distributions
from the Red Sea, the Indian Ocean and the western and central
Pacific to the Hawaiian Islands. An Indopacific species similar
to Pustularia is Nesiocypraea lisetae Kilburn,
1972 (fig. 1). It differs by having a distinctly curved aperture
and a distinctly denticulate fossula, which is narrow and smooth
in all Pustularia species. In adult shells of Nesiocypraea
lisetae there is a callous ridge connecing both, labral
and columellar teeth. In Pustularia, a comparable ridge
may be present on columellar side, especially in subadult specimens
of P. cicercula and P. margarita, but it is not
found towards the extremities as in N. lisetae. Adult
shells of all Pustularia species form a remarkable posterior
extremity composed of a callus-accumulation and two marginal
spines formed by the posterior terminal ridges (fig. 3). This
type of posterior extremity characterizes the Pustularia-shell.
It is not found in any other living Cypraeid species and also
lacks in the Caribbean fossil species assigned to Pustularia.
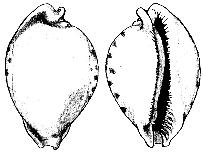
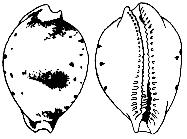
Fig. 1 (left): Proadusta moloni, Vicenza, Late Eocene
(20 mm), (right): Nesiocypraea lisetae, Richardsbay, Natal
(15 mm)
2. Comments on the
taxonomy of some living Pustularia
A great deal of confusion was
caused by Schilder (1966) concerning the type species of Pustularia,
P. cicercula (Linné, 1758). He studied the Linnean
collection in the British Museum in 1936 and based his later
descriptions of subspecies (Schilder & Schilder 1938) on
this experience, whereas the results from the 1936 visit were
published only thirty years later.
Linné hardly ever designated type specimens, and in many
cases it has been difficult to derive sufficient information
from his brief descriptions. The correct usage of names such
as labrolineata and flaveola is still being debated:
just two examples from the Cypraeidae.
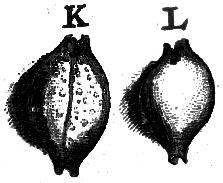
Fig. 2: Type figures from Rumphius (1705)
Plate 39. K: cicercula, L: margarita
Schilder did not find specimens of cicercula in the remains
of the Linnean collection, but he located two shells he identified
as bistrinotata bistrinotata: those were reported
to be badly corroded, without basal blotches, but with very faint
spots and blotches dorsally. There are traces of a dorsal groove.
Schilder concluded that these two shells were Linné's
types of cicercula, hence synonymous with bistrinotata.
He proposed to change the usage of the name cicercula
for the shell formerly known as bistrinotata. In the brief
Schilder description of cicercula, Linné does not
mention any dorsal blotches, but to Schilder there seemed no
doubt that the two eroded shells in the Linnean collection were
the types of cicercula.
Linné was a very accurate observer, and it therefore appears
unlikely that dorsal blotches would have escaped his attention
or were considered insignificant.
For many reasons, later authors did not accept the taxonomic
changes postulated by Schilder, and indeed they appear too forward.
himself admitted in the 1966 work: For though we cannot be absolutely
sure that the specimens preserved in Linnaeus' collection are
the same shells which Linnaeus faced when he composed his descriptions,
there is a probability in different degrees that it was so. Taking
this into account, it is confusing why Schilder & Schilder
described bistrinotata in 1937, only one year after studying
the questionable shells in the Linnean collection.
There is no explicit holotype specimen of cicercula. The
figures and the text of the original description are all we have
at hand. Linné, like many other later taxonomists, based
many of his taxa on illustrations from other publications, and
in the case of Mollusca, the work by Rumphius (1705) was a frequently
used source. A famous example is Conus aurisiacus Linné,
1758 which is depicted in perfection in this monumental work.
In the case of cicercula, the figure Linné based
his description on is also in Rumphius plate 39 fig. K (see also
Dodge 1953). This illustration shows a pustulose shell with a
distinct dorsal groove and a spire blotch. Apparently, this figure
in Rumphius shows a cicercula in the sense the name is
in use today, as the combination of a dorsal groove and a spire
blotch is specific. Schilder himself did not follow his own proposal
of 1966, as in the Catalogue of 1971 the traditional usage of
cicercula and bistrinotata is maintained (see also
Burgess 1977).
Incidentally, in the same work by Rumphius there is the well
recognizable original type figure of margarita (Dillwyn,
1817) (pl. 39 fig. L), explicitly described as being completely
smooth compared to the granulose shell (knorrelige Kopje) of
fig. K on the same plate (= cicercula). These illustrations
from Rumphius are reproduced here (Fig. 2).
In the above-mentioned 1966 article, Schilder described the supposed
types of globulus from the Linnean collection. These shells
show the characteristics of the typical globulus globulus
coming from Sri Lanka and Thailand and correspond with s description.
This is one case in which the assignment of a Linnean name is
referable to actual specimens in his collection and most likely
correct, even at the subspecific level.
The taxa introduced by Schilder & Schilder are: bistrinotata
bistrinotata (1937), b. mediocris, b. sublaevis,
and globulus sphaeridium (all 1938) and finally
bistrinotata keelingensis (1940). Their characteristics
and measurements, along with the type localities are compared
precisely in the Prodrome of a Monograph p. 126-127. The interpretation
of these names, as well as their correct assignment to the various
series of shells investigated therein presents no problem.
The recently described Pustularia jandeprezi Poppe &
Martin, 1997 will be discussed below.
Finally, mauiensis Burgess was based on a series of shells.
The species was illustrated and described and compared carefully
and accurately.
3. The conchological
features of Pustularia
The shells of Pustularia
can readily be separated from others in the family Cypraeidae
by the globular body, with the above-mentioned spiny-rostrated
extremities in combination with an extremely narrow aperture
framed by fine dentition. A pustulose surface is common. A second
ridge formed by the teeth on the columellar side is sometimes
found inside the aperture, while the edge of the moderately narrow
fossula is always smooth. The spire is slightly projecting in
the subadult, but completely concealed by callus in the adult
stage. The species of Pustularia differ from each other
mainly by different combinations of very few features, some of
which are not found in other Cypraeidae. The terminology for
the Pustularia shell is shown in Fig. 3.
The most important features for taxonomic distinction are the
presence or absence of
a) the spire blotch
b) dorsal granules
c) the four basal blotches
d) the three dorsal blotches
To judge the value of the various shell features, it proved necessary
to examine larger series of specimens from defined geographic
origins. The comparisons were made on series consisting of at
least ten specimens. Table 1 shows the results of this study
of 32 lots of more than 1100 specimens from 20 localities. Values
printed in bold letters are those of highest consistency, denoting
that 100% of the examined specimens of the respective lots show
the feature. Those entries printed in standard letters describe
lots in which 75% or more of the examined specimens show the
particular feature. The remark variable denotes that a feature
was found to be highly variable and hence of no value for distinction.
For the comparative analysis, a consistent feature (100%) is
an argument in favor of a distinction at the species level, while
in a subspecies or a forma, a certain number of qualitative (75%)
features may be sufficient.
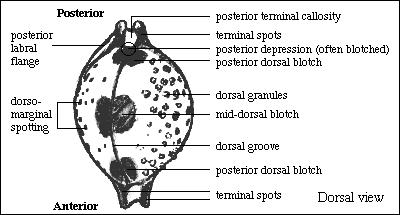
Fig. 3a: Terminology of the shell of Pustularia (dorsal
view)

Fig. 3b: Terminology of the shell of Pustularia (basal
view)
Explanation of the categories (features) in Table
1:
The lots and localities (in alphabetical order):
•And: Andaman Sea, a small lot of shells collected by divers
between 1991 and 1994.
•Eilat: Specimens collected between 1985 and 1990.
•Hawaii: Olavalu, Maui Is., collected in shallow water between
1983 and 1990 ( mauiensis), and fifteen specimens of a
cicercula-like population from Oahu (see chapter 5).
•Keel.: Cocos/Keeling Island, collected between 1967 and
1975.
•Kwa.: Kwajalein Atoll, a large lot of more than 200 specimens
collected by Scott Johnson in depths of 30-35 m between 1990
and 1996.
•Maurit: Mauritius, a lot of beach-collected material from
1981 to 1984.
•NCal.: Noumea, New Caledonia (1983-1986).
•Phil.: Philippines. The lot of P. chiapponii n.
sp. from Borongan, E. Samar Island, collected between 1985 and
1990. In addition to this lot of twelve specimens, there are
an additional 40 specimens supplied by the company Tridagna (Spain)
who could not give details about their origin except Philippines.
Those shells correspond exactly with those from ascertained origin
so that they were included in the study. The other lot marked
Philippines is from Samar Island, collected between 1979 and
1995.
•Taiwan: A lot of shells collected between 1990 and 1992
in the south of Taiwan.
•Tahiti: Specimens collected by divers in the area around
Hitiaa, Tahiti between 1986 and 1995.
•Thail.: Specimens from Phuket island, Thailand, collected
between 1985 and 1990.
•Tua.: Tuamotu, a lot of shells labeled Tuamotu ’69
from the Eva Roscoe collection.
•Tuléar: Madagascar, collected by snorkeling, 1996.
•Samoa: specimens from the northwest coast of Savaii Island,
Western Samoa, collected between 1992 and 1996.
•Solom.: Malaita and Marau Sound Islands, collected between
1965 and 1975.
•Sri La.: from Beruwala to Kosgoda Beach, S. W. Sri Lanka,
a larger lot of shells collected on the beach after storms, between
1990 and 1997.
•Zanz.: Zanzibar Island, a large lot of shells collected
on the shore after the cyclone of 1967.
Size: Obtained from the average measurement of all available
adult specimens.
General shape: Although shape in Pustularia is
rather similar in all species compared with other cypraeids,
the tendency of the shell’s body (not considering the extremities)
was compared.
Dorsal color: The ground color of the dorsum varies little
throughout the genus; it was found to range from cream to brown,
with variable shades of yellow or orange. Plain white shells
were found to be an exception.
Dorsal blotches: These actually consist of three pairs
of darker staining on either side of the dorsal line (the name
bis-tri-notata refers to these). The mid-dorsal pair is often
missing while traces of the anterior dorsal blotch are rather
common. The complete absence of any trace of dorsal blotches
safely separates a number of species from those groups which
occasionally have such traces. Caution: In P. cicercula,
for instance, there may be an accumulation of brown pattern along
the dorsal line, but this is not the same feature, as it is formed
by the secondary (adult) pattern. The three pairs of dorsal blotches
described here are already formed by the subadult shell (see
below), they are always absent in P. cicercula.
Terminal spots: Some species show a darker staining of
the extremities visible only in the dorsal view. In Pustularia,
this feature seems of little importance for species and subspecies
characterization.
Dorso-marginal spots: The marginal spotting in Pustularia
species usually extends also onto the dorsum, where it mixes
with a very fine brown mottling or spotting. The latter is a
very variable feature and was not compared, while the coarser
spotting itself was found to be of relative importance. In granulose
species, a spot usually occurs together with a granule.
Basal blotches: In some Pustularia species there are
two marked blotches on either side of the aperture. This feature
was found to be of considerable importance for separation of
species and subspecies. In some taxa, the blotches may be small,
incomplete, or faint, and sometimes present only in a certain
percentage of specimens. In those cases a statistical comparison
may be important for identification, especially on the subspecific
level.
Dorsal groove: This feature is usually correlated with the
degree of dorsal granulation. Heavily granulose specimens usually
have a well-defined and deeply cut dorsal groove. This feature
is important for separating species in those cases where it is
never found vs. those taxa where it occurs at least occasionally.
Blotch at spire: Even if a dorsal groove does not exist
there is a more or less distinct depression left of the spire
in adult shells. In two species, this depression is distinctly
blotched darker while it is not in all other taxa.
Dorsal granules: The degree of granulation and the areas
of the dorsum showing granules may vary considerably. The mere
fact whether granules may ever be found or not is important for
species distinction. The taxa compared in Table 1 are thus interpreted
as either granulose (distinct, indistinct or variable) or smooth
(absent).
Aperture: The aperture of all Pustularia is very
narrow and of the same width throughout. In most taxa, it is
rather straight or very slightly curved to the left. There are
some taxa, however, in which the aperture shows a distinct curve
to the left in the posterior third of the shell.
Shape of extremities: Although in all species of the genus,
the extremities have about the same structure, the degree of
rostration and the thickness of the terminal spines varies. The
data in the table reflect the general impression gained from
comparison of the various series.
Dental color: Darker staining of the teeth usually varies
considerably within a single lot, but there are certain groups
in which this feature is highly consistent. Comparison of the
dental color is always made with the color of the middle of the
base.
Average number of labral and columellar teeth: The teeth
of each specimen of the lot were counted excluding indistinct
crenulations towards the extremities. The numbers given here
are the actual number of teeth counted, not the normalized number
after Schilder & Schilder 1938 (relative number of teeth
in a hypothetical shell of 25 mm length).
Length of teeth and columellar callus bridge: The
length of the teeth is described as short = not reaching the
middle of the labrum or mid-base, medium = reaching onto the
middle of the labrum or base, or long = reaching far across the
labrum and base, touching the margins. In some shells, there
is an accumulation of callus more or less covering the teeth.
This cover may be present only along the aperture, forming a
narrow bridge across the teeth. In some cases, the callus is
massive and covers all of the teeth in the mid-third along the
aperture.
The data in Table 1 reveal that at least five species can be
defined on the basis of consistent features. It is also apparent
that a population of aberrant shells was examined whose conchological
relationship to bistrinotata is obvious, apart from very
constant features that suggest a distinction at the species level.
This population is described below as Pustularia chiapponii
n. sp.
4. A new species of
Pustularia from the Philippines
Material
Two lots of live-collected shells were obtained recently from
commercial shell dealers. One lot of twelve specimens is from
Borongan, Samar, and the other lot of 40 specimens reached the
author without locality data except Philippines. Both lots were
mixed with larger quantities of cicercula, margarita
and globulus, as well as typical bistrinotata bistrinotata,
suggesting that the origin is indeed the Philippines. Since both
lots of shells show identical shell features, it is assumed that
they were collected in the same area. It is astonishing that
despite the vast exploitation of the Philippines Islands, even
today shells appear which have never before been illustrated
or studied. The new taxon is described here in honor of Dr. Marco
Chiapponi of Lecco, who has contributed for a long time to the
study of the interesting genus Pustularia.
Pustularia chiapponii n. sp.
Description
Greatly inflated, globular. The extremities are distinctly rostrated
but barely margined. The spire is large, somewhat projecting,
completely covered by a thick callus. Dorsum and margins are
densely granulose; those granules are less distinct towards the
middle of the dorsum. There is a deep dorsal groove running longitudinally
across the shell, forming a Y-shaped depression bordering the
posterior extremity. The posterior terminal callosity is well
developed. The aperture is narrow and conspicuously curved to
the left in the posterior third.
The teeth on both sides are fine, rather weak, but well defined
throughout their way across both labrum and base and far onto
the margins on both sides where they meet the dorsal pustules.
The posterior labral margin is crossed by the teeth, which thus
are visible as spiny processes in the dorsal view. The entire
shell is pale yellow, there are no traces of basal blotches,
nor is there any staining of the teeth. The yellow color is slightly
paler only towards the aperture. There are three barely perceptible
darker patches dorsally, which may be interpreted as blotches.
These are, however, not separated by the dorsal groove. There
are no traces of smaller marginal or dorsal spots.
The paratypes correspond exactly with the holotype, none of the
features described above are subject to perceptible variation.
Designated types
Length x width x height (mm), (labral teeth: columellar teeth),
depository
Holotype: 19.1 x 12.6 x 11.4 (24 : 22) coll. HDN Cismar
Paratype 1: 19.1 x 12.6 x 11.6 (24 : 18) coll. Chiapponi
Paratype 2: 16.4 x 10.4 x 9.4 (25 : 18) coll. Chiapponi
Paratype 3: 17.7 x 11.5 x 10.2 (24 : 18) coll. Lorenz
Paratype 4: 19.4 x 12.3 x 11.0 (22 : 18) coll. Lorenz
Paratype 5: 18.7 x 11.5 x 10.3 (25 : 19) coll. Lorenz
Paratype 6: 19.7 x 12.3 x 11.0 (25 : 19) coll. Chiapponi
Paratype 7: 17.9 x 11.1 x 9.9 (24 : 18) coll. Chiapponi
Type locality
The only ascertained locality of the new species is Borongan,
Eastern Samar Island, Philippines. The habitat is unknown but
suspected to be rather shallow water.
Discussion
Pustularia chiapponii belongs to the group of pustulose
species without a spire blotch. The only similar species, therefore,
is P. bistrinotata from which it differs by the following
features: complete absence of basal blotches in all specimens
known. There are some specimens of bistrinotata, especially
of its Melanesian subspecies mediocris, in which the basal
blotches are reduced, incomplete, or even absent. In mediocris,
the percentage of specimens without basal blotches is 40% for
Solomon populations and 25 % for New Caledonian populations.
In 60% of the Solomon specimens and 75% of the New Caledonian
specimens, there are at least apparent remnants of basal blotches.
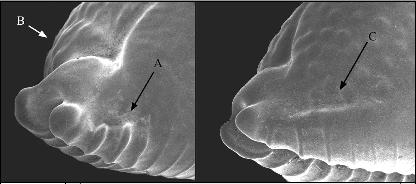
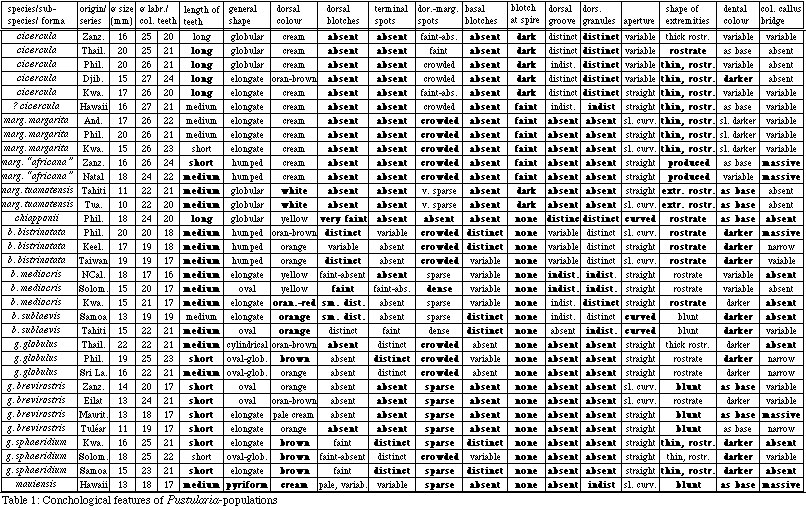
Fig. 4: The characteristic posterior extremity
of a Pustularia. Left: P. chiapponii n. sp., right:
P. bistrinotata. (SEM photos)
In chiapponii, the teeth are long, extending far across
the base and the labrum, they are never interrupted by a callus-bridge,
and in no specimen are they shorter mid-way on either side. In
none of the populations of bistrinotata are there teeth
of comparable length. Furthermore, the teeth meet with the dorsal
granules, which are often connected.
In P. bistrinotata, the pustules are usually separate
and distinct from each other (Fig. 5). The posterior labral teeth
cross the terminal margin (Fig. 4, left). This feature is seen
only in exceptional specimens of bistrinotata. All populations
of bistrinotata show a brown spotting of the margins as
well as the dorsum. Smaller spotting is not developed in chiapponii.
The dorsal blotches characteristic of bistrinotata are
almost absent in chiapponii, in which the darker zone
is not split in two by the groove mid-dorsally. The spire of
chiapponii is large, situated right of the posterior extremity,
and despite being covered by callus, is still visible as a slight
projection (Fig. 4, right). This is usually not seen in bistrinotata.
The distinctly curved aperture is typical in the Polynesian sublaevis
but rarely in populations of the Western Pacific. Finally, the
general shape of chiapponii differs by being more globular,
with the extremities being coarser and rather knobby.
The pale yellow color all over is only found in southeastern
Melanesian populations of mediocris, and especially in
New Caledonian shells. Those, however, are less inflated, their
dorsal groove is indistinct, the basal blotches are faint but
visible, the teeth are coarser and much shorter, and the posterior
columellar teeth do not cross the posterior labral margin towards
the dorsum.
A distinction of chiapponii from other members of the
genus represents no problem.

Fig. 5: The characteristic posterior extremity of a Pustularia
(columellar view) Left: P. chiapponii n. sp., right: P.
bistrinotata. (SEM photos). Note the connected pustules in
P. chiapponii.
Remarks:
1) In the meantime, three fresh dead and one living specimens
of P. chiapponii were collected by a diver from 25 m depth
at Divinubo Island, just opposite Borongan Bay, Samar. The specimens
were collected at night, lying upside down on the floor of a
small cave. They had probably dropped from the roof of the cave,
which was reported to be densely overgrown with sponges and soft
corals.
2) It is probable that on the international shell market, the
species will not be thoroughly recognized as it is easily confused
with new Caledonian bistrinotata mediocris. To summarize again
the main differences: in chiapponii, the teeth are more
numerous, and longer, reaching far across base and margins. The
pustules in chiapponii are finer and denser, and there
are no distinct darker spots or basal blotches. The aperture
is more corved, the extremities thicker. Click
here for a closeup comparison.
5. The living species
of Pustularia
A cladographic comparison of all
species and subspecies of the genus based on those features whose
statistic consistency is greatest would produce relationships
given in Fig. 6.
According to this interpretation, the presence or absence of
a spire blotch is ranked as a conservative feature that has been
lost completely in one lineage while it was retained in the other.
It is the most consistent and apparent feature, along with a
number of linked characteristics separating the cicercula/margarita
group from the globulus/bistrinotata group in which
basal blotches and orange to red ground color may occur.
The dorsal granulation was reduced independently (as a secondary
adaptation) in both lineages: in the cicercula/margarita group
( margarita) and in the globulus/bistrinotata group
( globulus, and to a high degree in mauiensis).
It is a feature shared with the related Indo-Pacific Staphylaea,
but not with the older ancestral Atlantic forms, such as Proadusta.
Fig. 6: A possible cladographic interpretation of the living
species of Pustularia
In mauiensis, rudimentary granulation is present in occasional
specimens, as is the basal and dorsal blotching. This places
mauiensis in close proximity to bistrinotata. In
all three, globulus, bistrinotata, and mauiensis,
the basal blotching is either well-produced or at least rudimentary,
whereas in chiapponii it is lost.
The number of features that are not found in chiapponii
(blotches on base and dorsum, marginal spotting, no columellar
callus-bridge) weigh against those which are specific for chiapponii
(longer teeth, connecting granules, curved aperture, projecting
spire, denticulate posterior labral margin). The features compared
in the cladogram are merely of a conchological nature and do
not necessarily reveal biological relationships. Their evaluation
in the cladogram shown here is not the only possible one, but
a consequence of the above-made interpretations.
P. cicercula and margarita form a group of two
closely related taxa separable by one consistent conchological
feature only. Both have a distinct spire blotch that is not found
in the other taxa. Furthermore, both have no traces of basal
or dorsal blotches. Recent radula studies confirm their status
(Lorenz, 1998). Bradner & Kay (1996) also illustrate the
radulae of cicercula and margarita. Their illustrations
show radulae of certain variability but with apparent differences
between the taxa.
In specimens from Kwajalein, the mantle of cicercula shows
a darker spotting, the papillae are rather thin and short. In
margarita from the same locality, the mantle is translucent,
with only few white spots encircling the somewhat thicker and
longer papillae. How well these features can be used to draw
conclusions, however, is so far uncertain. Interestingly, the
somewhat rarer margarita has a wider distribution and
a far greater geographic variability compared to cicercula.
Three forms are compared in the table. The rare western africana
hardly differs from typical margarita, whereas the features
of tuamotensis are rather constant and remarkable. It
is suggested that tuamotensis deserves subspecific rank.
As can be seen by comparing the data in Table 1, P. cicercula
shows little variation throughout its wide distribution. Specimens
from the western Indian Ocean are occasionally distinguished
as subspecies or form lienardi (Jousseaume 1874). The
features assigned to this taxon by Schilder & Schilder 1952
simply cannot be ascertained for East African or Red Sea specimens,
merely the smaller average size is relatively constant.
A set of shells from Oahu, Hawaii should be mentioned here, provisionally
encorporated as ? cicercula in Table 1. Conchologically,
these specimens have intermediate features between cicercula
and margarita, as the dorsum is usually smooth on top
but faintly pustulate towards the rostrated extremities. The
teeth are very fine and unstained. The most striking feature
is the reduction of the spire blotch, which is barely visible
in some specimens.The systematic position of this population
is so far under study.
The group of shells without a spire blotch is split into four
species:
P. chiapponii, in which no basal blotches, no marginal
spotting, but a distinct granulation are found. It is the least
variable species with a very limited distribution in the Philippines.
P. bistrinotata the most variable species, with a number
of geographic subspecies and varieties, which are compared in
Table 1. Generally, the species is characterized by the combination
of four basal blotches, three pairs of dorsal blotches separated
by a more or less distinct sulcus, and the pustulose surface.
In the western b. bistrinotata the dorsum is rather humped,
the teeth are nearly always stained darker, the basal blotches
are distinct and large, the dorsal blotches are dark and wide,
the dorsal ground color is a saturated reddish-brown and the
marginal spots are dense and dark.
The name keelingensis was introduced for specimens from
the Indian Ocean. It is described as being more humped, less
granulose, without distinct dorsal sulcus and less produced teeth,
the basal blotches are described as being well-marked. Such shells
are found throughout the Philippines, and mainly Samar and Bohol,
but the concept of keelingensis was not found to characterize
particular specimens from Cocos/Keeling Island and other western
localities. It is considered a syynonym of bistrinotata bistrinotata.
In Melanesia and Micronesia, the paler and more elongate mediocris
occurs. In Micronesian populations the shells are rather small,
rostrated, and bright orange, while Melanesian specimens are
usually yellowish. Callused, larger and rather pale New Caledonian
specimens have been separated as forma cicerculaeformis
Raybaudi, 1993.
The less granulose, more distinctly curved and blotched Polynesian
sublaevis is considered a subspecies here, judging from
the series examined. However, the separation from mediocris
is sometimes difficult and only possible when larger numbers
of shells from ascertained localities are compared.
As already mentioned, the Hawaiian P. mauiensis occasionally
shows two or even all four basal blotches and, more commonly,
three dorsal blotches. This phenomenon reveals a close relationship
with bistrinotata. In many specimens a very indistinct
granulation is visible on the anterior portion of the dorsum.
The status of mauiensis as a valid species can be confirmed
by this study, mainly because of the coarser, shorter and always
unstained teeth, as well as a different dorsal and basal ground
color. The basally blotched populations from northern Oahu have
been separated as ssp. wattsi. DNA analysis confirms their
status as separate entity.
P. globulus seems subdivided into four geographical subspecies:
The western brevirostris Schilder & Schilder, 1938
is characterized by smaller size, absence of staining on the
teeth, and lack of basal blotches. It is a well-distinguished
geographic subspecies ranging from the East African coast to
Mozambique, the Seychelles to the Mascarenes, maintaining the
characteristic features.
Specimens from the Red Sea show a dark staining of the teeth,
which suggests that (despite smaller size) these belong to g.
globulus rather than brevirostris. They have been
described as g. giftunensis on the basis of Pleistocene
fossils. That name is now adopted for the living Red Sea globulus.
The shells of g. globulus from the
northern and Central Indian Ocean are often cylindrical rather
than globular, the teeth are only faintly marked with brown,
the extremities are short and rather solid, four basal spots
are rarely seen.
Smaller, more rostrated shells with darker teeth and four distinct
basal spots are found in Pacific regions, especially Melanesia
and western Polynesia. This eastern form is well known as P.
globulus sphaeridium Schilder & Schilder, 1938. The development
of typical globulus to sphaeridium is rather gradual,
as can be seen from the data in the table.
Philippine specimens may already resemble those from the Solomons
which can be assigned to sphaeridium. Still, the status
of sphaeridium might be considered as a geographical subspecies
with considerable overlapping zone in the western Pacific, while
the populations from western Polynesia and Micronesia are consistent
in their features and have little resemblance to classic specimens
from Sri Lanka or Thailand.
The systematics of the living members of the genus Pustularia
according to this study looks as follows:
Genus: Pustularia
1. cicercula (Linné, 1758)
syn.: lienardi Jouss. 1874, vulavula Steadm. &
Cott. 1943
2.1 margarita (Dillwyn, 1817)
syn.: tricornis Jouss. 1874
2.2 margarita tuamotensis Lorenz & Hubert,
1993
3.1. bistrinotata bistrinotata Schilder & Schilder,
1937
syn.: quadrimaculata Dautz. & Bouge 1933, keelingensis
Sch. & Sch. 1940, jandeprezi Poppe& Martin, 1997
3.2. bistrinotata mediocris Schilder & Schilder, 1938
3.3. bistrinotata sublaevis Schilder & Schilder, 1938
4.1. globulus globulus (Linné, 1758)
syn.: affinis Gm. 1791, tetsuakii Kira 1959
4.2. globulus brevirostris Schilder & Schilder, 1938
4.3. globulus sphaeridium Schilder & Schilder, 1938
4.4. globulus giftunensis Lorenz 1992
5.1 mauiensis mauiensis (Burgess, 1967)
5.2. maueinsis wattsi Lorenz 2001
6. chiapponii Lorenz 1999
6. The subadult shells
of Pustularia
Subadult bistrinotata show
three blurred darker basal blotches, which later become encircled
by darker spotting rising upward from the margins. The basal
blotches form rather late in development, as do the dorsal granules.
In general, subadult shells of Pustularia have shorter
teeth, a high gloss, no development of granules, a partly exposed
spire and a posterior extremity without the characteristic structure
seen in all adult Pustularia.
Recently, P. jandeprezi Poppe & Martin, 1997 was described
from the Philippines and compared with bistrinotata. First
impression of the diagnosis suggests that it is based entirely
on subadult specimens of bistrinotata. In fact, subadult
specimens of bistrinotata are found throughout its distribution,
and they do resemble the holotype of jandeprezi in every
respect. The authors explain that their type is a fully adult
shell with well-formed dentition and extremities and a callused
base.
 |
Pustularia jandeprezi (bistrinotata sudadult ?) Philippines
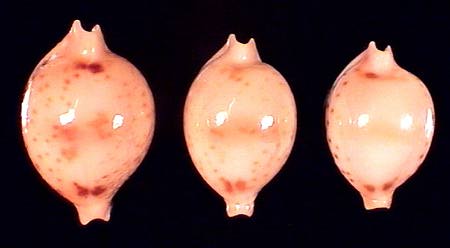 |
Pustularia jandeprezi (bistrinotata mediocris sudadult ?) Solomons
The description reads as follows:
Shell thin, solid, very globose. The general shape is oval-globular.
The extremities are thin, solid and long. The teeth are fine
and short. The callus on the base is thick, white to slightly
olive colored. Basal blotches may be present or absent. The dorsal
color varies from cream-white to olive-brown, most often with
a mid-dorsal blotch. The rounded dorsum is smooth and no specimens
with dorsal granules are known. The teeth on the columellar side
number between 20 and 25. The labial teeth between 21 and 28.
The spire is prominent, even in heavy adults. A dorsal line is
absent. One paratype [No. 11] is almost entirely white colored,
without marginal spotting, and with a pale mid-dorsal
blotch (...) [I suspect that latter specimen, which shows
absolutely no similarity to any of the other types might be P.
chiapponii].
In the discussion, Poppe and Martin stress the absence of any
dorsal granules and color differences compared to bistrinotata
from the same area, as well as the exposed spire even in callused
shells of jandeprezi. The latter is always concealed in
adult bistrinotata. There is, according to the authors,
also a difference in shape and number of teeth compared to bistrinotata.
Comparison made between subadult specimens of bistrinotata
from various localities with the illustrations of the jandeprezi
types did not reveal any significant difference, but as the
holotype is claimed to be an adult shell, the status of jandeprezi
remains an open question.

Fig. 7: Pustularia margarita, subadult.
SEM-Photograph (500x)
Sudadult shells of other Pustularia species
are translucent and mostly white. The Western brevirostris
is plain white, the extremities short and spiny. On the columellar
side, a longitudinal denticulate ridge forms prior to the strong
dentition, which develops as the shell reaches maturity. Also
in cicercula and margarita, the young shells are
white, translucent, with a comparable columellar ridge. The latter
is typical in adult specimens of the rare Nesiocypraea
lisetae, for which subadult cicercula or margarita
are occasionally mistaken by dealers. In early stages of all
Pustularia, the spire is still exposed. In such shells,
the surface (especially on labral side) shows longitudinal growth
ridges visible under magnification (Fig. 7). In cicercula
and margarita, the subadult shells show two darker transverse
bands that are never preserved in fully adult shells.
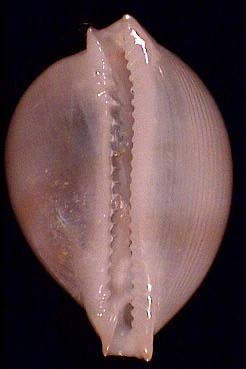 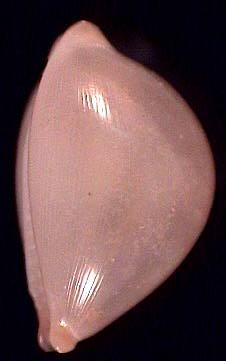 |
Pustularia margarita (sudadult), Philippines
7. Relation between
locality and average shell length
Another interesting aspect that can
be derived from Table 1 is the change in average size from western
to eastern localities. Splitting the Indo-Pacific into five sectors,
the average shell lengths of each sector vary considerably within
a species: shells from the Western Pacific are largest, while
those from East Africa and eastern Polynesia are considerably
smaller (Fig. 8).
The reasons for this phenomenon are so far unknown. In other
genera of Cypraeidae a similar increase in size from western
to eastern localities has so far not been confirmed.
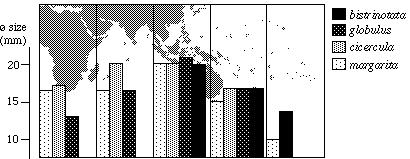
Fig. 8: Change of the average shell size with geographic region
8. Colour plates
Acknowledgements
The present study could not have been
carried out without the assistance of colleagues and friends.
Many thanks to
Prof. Dr. Alex Hubert †, Scott Johnson (Kwajalein) Prof.
Dr. Dr. em. Günther Sterba (Markkleeberg); Dr. Hans Werner
Koyro (Gießen); Prof. Dr. Klaus-Jürgen Götting
(Pohlheim); Prof. Dr. Erhard Schulte (Alten-Buseck), Mohammed
Makungu (Zanzibar); Guido Poppe (Berchem); Dr. Marco Chiapponi
(Lecco), Ludwig Gabrielli (Neuss), Dr. Vollrath Wiese (Cismar),
Xavier Pallarols Blanch, Capt. Felix and Ingrid Lorenz (Lauenburg),
and Jana Kratzsch (Buseck-Beuern).
Literature
Burgess, C. M. (1967): A New Hawaiian Cypraea The Nautilus
81:6.
Burgess, C. M. (1977): Here's more about Cypraea labrolineata
Hawaiian Shell News 207
Burgess, C. M. (1985): Cowries of the World Cape Town, Gordon
Verhoef, Seacomber Publications.
Bradner, H. & Kay, A. (1996):An Atlas of Cowrie Radulae (Mollusca:
Gastropoda: Cypraeoidea: Cypraeidae) The Festivus XXVIII Supplement.
Dillwyn, L. W. (1817): Descriptive Catalogue of Recent Shells
London, 1: 470
Dodge, H. (1953): Mollusks of Linnaeus Bulletin of the American
Museum of Natural History 103:118-121
Lorenz, F. and A. Hubert (1993): A Guide to Worldwide Cowries.
Christa Hemmen Verlag, Wiesbaden.
Lorenz, F. (1998): Kauris von Ostafrika. Schriften zur Malakozoologie
11, 1-150.
Poppe, G. T. & Martin, P. (1997): A New species of Pustularia
from the Philippines (Gastropoda: Cypraeidae) Gloria Maris 35
(6) 81-88.
Raybaudi, L. (1993): Genus Pustularia World Shells 5:68.
Rumphius (Rumph, G. E.) (1705): Amboinsche Rareitkamer Amsterdam:
113.
Schilder, F. A. & M. Schilder (1938): Prodrome of a Monograph
on Living Cypraeidae. Proceedings of the Malacological Society
London 23: 119 231.
Schilder, F. A. & M. Schilder (1952): Ph. Dautzenbergs Collection
of Cypræidæ Mémoires Institut Royal des Sciences
Naturelles de Belgique (2. ser.) 45: 1243 and Pl. IIV.
Schilder, F. A. (1966): Linnaeus type specimens
of cowries The Veliger 9: 91100.
Schilder, F. A. & M. Schilder (1971): A catalogue of living
and fossil cowries Mem. Inst. Royal Sci. Natur. de Belgique 85
(1971) 1246.