
Parasitic Mollusca
On some families with parasitic life-strategies in the phylum Mollusca
Modern parasitology can be divided in three mainstream disciplines,
each of which have their starting point from the hosts involved: the
first has a medical
approach as it dwells upon those types of parasitism causing disease in
humans. The others study those types of parasitism causing economical
damage, that is disease to domestic animals and plants. The parasites
involved are either Protozoans, worms in the widest sense
(Plathelminthes, Nemathelminthes), and certain Arthropod families.
While there is a wide range of literature dealing with the
life-strategies of these groups, no monographic work has ever been
published on the Mollusca as a phylum bearing a wide range of parasitic
species and an even wider range of strategies.
The phylum Mollusca consists of approximately 150.000 species. There are eight classes:
Caudofoedata and
Solenogastres (these two are also known as Worm-Molluscs)
Polyplacophora (Chitons)
Monoplacophora (a small group of archaic deep water snails of limpet-like shape)
Gastropoda (Snails)
Bivalves (Mussels)
Scaphopoda (Elephant-Tusks)
Cephalopoda (Squids and Nautilus).
The majority of species belong to the Gastropoda and the Bivalvia.
These two classes are also those with a wide range of parasites.
The term parasitism is derived from the greek para (next to...) and the latin situs
(site, position), referring to favourites lying next to their sponsors
on grand dinner banquets, taking advantage from a position right beside
their host. The latin word parasitus has two meanings: the table-guest, and the parasite.
Parasitism refers to a variety of life-strategies which may have developed differently. The parasite is the one taking advantage from the host, whose disadvantage defines the relationship as parasitism. Those relationships which are to the advantage of both are called symbiosis.
Many types of parasitism have developed from symbiosis, whenever one of
the partners would start to take profit to the discadvantage of the
other.
Another relationship called phoresis occurs when an animal or
plant uses another species as a mean of transportation. It is mostly
temporary and not to the disadvantage of the "taxi". Parasitism may be
temporary or permanent, it may be closely linked to a particular type
of host (obligatory), one or many hosts may be involved. We are
speaking of endoparasitism when the parasite lives permanently inside the host, or ectoparasitism,
when the parasite sits somewhere outside, on the surface of the host.
There are temporary types of parasitism called brood-parasitism, when
the host is used only for the larval development of a species. An
extreme case is the so called nurse-eggs, in which only one larva
develops while eating all its sisters and brothers in the same capsule
(larval cannibalism).
Most real-life types of parasitism are mixtures or combinations of
these various theoretical models, and examples for all of them are
found in the Mollusca. In the following, a few examples will be shown.
Predators on sponges
The most primitive type of parasitism is probably that performed by
many molluscs feeding on sponges. The host is not killed (or eaten up)
completely, but suffers severe damage. The ancient group of slit shells
(Pleurotomariidae) all feed on sponges but may also be found feeding on
crinoids and other animals. The parasite/host relationship is rather
loose.

Perotrochus midas feeding on a sponge
In the Cowries (Cypraeidae) a number of genera feed on sponges, too. In
some cases, the relationship to the sponge as food and shelter has
become obligatory. In Zoila marginata an interesting transition between the southern and the northwestern subspecies can be observed. Whereas the southern marginata feeds on smaller sponges and also moves from sponge to sponge, the northwestern ketyana permanently lives on one sponge and eats cavities into its "tissue".

Zoila marginata marginata feeding on a small sponge

Zoila marginata ketyana eating itself into a sponge
On the photo it is apparent that the sponge forms a cavity around the
posterior portion of the shell, hiding the produced lateral flanges
typical in this subspecies. Single specimens of ketyana
were reported to have eaten a tunnel deep into a sponge, into which the
animal could retreat for shelter. This is the first step from a
predatory animal to a parasite living in an association with a specific
host.
In the South African Cypraeovula mikeharti another step towards endoparasitism may be observed. Some individuals live under and inside the black sponge Guitarra. Also the egg-capsules are placed into cavities of the host. The closely related Cypraeovula algoensis from the same geographic area merely feeds on sponges, but the relation to the host is less obligatory.

Cypraeovula mikeharti
Parasites on Tunicates
The species of the Triviidae and Lamellariidae comprise many species of
obligatory parasites whose life-cycle is closely linked with a
tunicate. In the genus Trivia,
the female posesses a tube like structure called ovipositor. With the
aid of this orgen, egg capsules are deposited into the openings of
commensal tunicates. The larvae find shelter and food in their host.
Adult Triviidae may feed on sponges, algae and also tunicates.
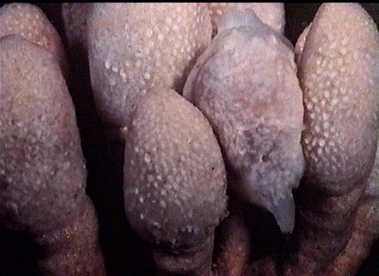
Trivia magnidentata in a colony of tunicates. The mantle perfectly camouflages the parasite animal
Parasites on and in corals
Most species of parasitic molluscs are found in association with
corals. The family Ovulidae consists of hundreds of species with a
different degree of host-specificy and various transitional stages from
commensalism, via brood-parasitism and host-specific parasitism.

Simnia spelta on a host coral
But there are also transitions away from Parasitism to Symbiosis. This
is very interesting, as the evolutionary way that has lead into
parasitism is postulated to be irreversible by many biologists. A group
of animals that has evolved into parasites has undergone a variety of
morphological adaptations to the host organism. A reverse evolution
appears unlikely. In Pedicularia
this seems to be the case, however. The group has evolved from
parasitic Ovulids some twenty million years ago and now occurs with a
handful of species on stylasterine corals in all temperate seas. The
shells are modified individually to fit exactly to one spot on the
coral, which over the years forms a cavity for the mollusc and its
spawn. The shell cannot move from this position but has a long
proboscis to reach far around. The former parasite no longer feeds on
the polyps of the coral itself, but on the mucus that is secreted by
the polyps, subsequently "cleaning" the colony.

Pedicularia sp. on a stylasterine coral
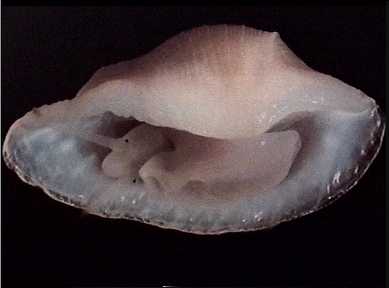
Pedicularia elegantissima, living animal, removed from host
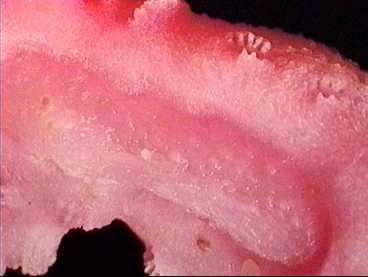
Cavity in the coral where a Pedicularia was sitting.
In the large family of the Coralliophilidae there are many examples of
coral-parasites, some of which are well known for their most bizarre
shells. The endoparasitic species are living more or less buried in the
tissue of the coral. Some anthozoan colonies were reported to grow
larger and denser when a particular Coralliophilid is present. Such
cases suggest that also here the step from parasitism towards symbiosis
was performed, as the anthozoans of such colonies were all undamaged.
Also in these cases, the Coralliophilid seems to feed only on the
anthozoan secretions, which are rich in protein and saccharids.

Rapa sp. in a soft coral

The same individual, removed from its host
Parasites on and in Echinoderms
There are many groups of ectoparasitic snails on starfish and Sea-urchins. Most of these belong to the Opisthobranchia, such as Ophioderma, a small snail that lives on the oral area and the arms of ophiurids, with their proboscis eaten into the tissue of the host.
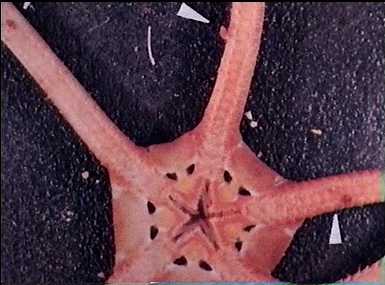
Ophioderma sp. on its host
Other limpet like shells (Neritina) live half-buried in the arms of the blue Linckia-starfish.
But there are also species of uncertain position that live inside the tissue of Sea-cucumber. The veligers of the genera Enterocolax and Enteroxenon
attach themselves to the skin of their host and actively drill into the
flesh, leaving their shells. The adult parasite is hardly separable
from the tissue of the host. These extreme cases of endoparasitism are
poorly investigated, but they show that some molluscs have found ways
to this highest level of parasitism.
Parasitism on and in fishes
The larvae of fresh water mussels face the problem that the current
would carry them into the sea. The populations would be swept out of
the rivers. The problem was solved by using fish as vector against the
current. The glochidian-larvae of almost all freshwater mussels spend
the time of their development-circle in the gills of fish moving them
upstream. The European freshwater pearl-mussel Margaritifera is dependent on the presence of the river-trout Salmo trutta fario. The introduction of the larger, faster growing and easy to breed rainbow-trout Oncorhynchus mykiss
to European rivers and lakes, as well as pollution has depleated the
populations of the river-trout and brought the pearl-mussel near
extinction.
But there is also another bizarre kind of parasitism that was
discovered only a few years ago, involwing parrot fish and the snails
of the buccinid genus Colubraria.
For a long time it was unknown what these shells ate as they have no
radula but a very long proboscis. This is one of the spectacular photos
of a sleeping parrot fish being stung into its mouth by Colubraria obscura, now known as the "vampire-snail".

- Colubraria obscura stinging a sleeping parrot-fish

The Cystiscid Kogomea sucking blood above the fin of a parrotfish. Photo Courtesy Scott Johnson
Parasitism among relatives- "nurse-eggs"
As mentioned above, many gastropods whose larval development lacks a
free swimming stage perform an interesting and apparently effective
strategy. A single egg capsule contains many hundred larvae, only one
of which develops further, eating the others in the capsule.
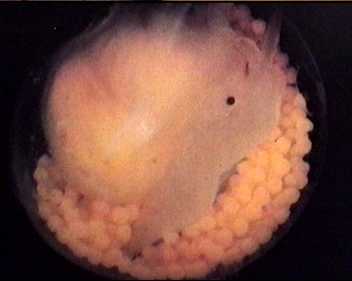
An egg-capsule of Cypraeovula algoensis. Only one specimen has developed to the pediveliger stage.
In the Plathelminthes the tendency towards parasitism is interpreted as
a strategy to compensate the lack of a protective shell or cuticula.
The main function of the host is to find shelter from outside
influences. In the Nemathelminthes the protective, acid-resistant
cuticula has encouraged parasitism simply because Nematods dont seem to
care in what medium they are as long as there is a food supply. In the
Arthropod groups with parasitic forms there are two aspects that have
supported parasitic lifes. First, the development of most effective
tools to suck, sting and bite, and second, the extreme selective
pressure by the success of this group, and the resulting urge to find
new ecological niches.
In the Mollusca, three main factors have lead to parasitism. First, to overcome the problem of immobility
and the consequenting danger of populations being swept out of rivers
(freshwater mussels). This immobility might have lead to phoretic
stages that later lead to parasitism. It is probable that most
parasitic relationships with Echinoderms result from phoresis. Second,
the pressure to find ecological niches in the densely populated marine biocoenosis. And third, the potential to be parasitic
by having developed organs variable enough to gather all kinds of food
and by body-shapes capable of hiding amongst- and clinging to- the
potential hosts.
This was uploaded in 2001. Subsequently I was asked to write a book chapter on Mollusca in: Marine Parasitology (2005) Csiro Publishing, Melbourne, p. 240ff