Notocypraea revisited The confusing group of Notocypraea from the southern half of Australia consists of a variety of taxa whose morphological features are highly variable and often inconsistent. Many authors have avoided dealing with that group and most collectors have simply "given up". The present taxonomic situation, proposed by Alex Hubert and myself and slightly modified in the process of study is the following (click the names for picture):
| species | subspecies | variations |
| pulicaria (+) | ||
| candida (-) |
||
| comptonii (+) | ||
| griffithi (-) | ||
| mayi (-) | ||
| casta (-) | ||
| ereginae (!) |
||
| piperita (+) | ||
| bicolor (-) | ||
| wilkinsi (-) |
||
| occidentalis (#) | ||
| euclia (!) |
||
| declivis (+) | ||
| dennyorum (#) |
||
| angustata (+) | ||
| verconis (#) |
||
| subcarnea (+) | ||
| dissecta (+) |
This classification is based on the following aspects: consistent conchological features (+), geographical (#), and bathymetrical features supported by fairly consistent conchological features (!), and finally, for the variation-status: slight conchological features not supported by geographical features (-)
The systematic arrangement of the genus now considers also genetical differences. These are rather slight in most cases, but surprising in others (e.g. verconis and dennyorum). The evolution of the shallow water taxa goes back to the Pleistocene age some 20 to 30 thousand years ago, when the ocean levels rose and covered vast areas of Australian land. These new habitats offered the chance for settlement and at the same time, presented a battlefield of different species competing for food sources, hiding places, ect. Random variability within each immigrant population and natural selection has lead to the formation and manifestation of new features, varieties and finally, species and subspecies, in the shallow water. The situation in the Notocypraea is comparable to that of the genus Zoila. The migration into the shallow water habitats that formed with the rise of the ocean-levels was slow, and it was more of an up- and -down rather than a gradual one. The archaic ancestral populations from which the shallow water immigrants originated may have looked pretty much the same as those forms we can find in greater depths today. In the deep water, the ecological conditions have hardly changed at the same time, the populations could remain more stable and retain more archaic features. Molecular data confirms that they are archaic (dissecta and subcarnea).
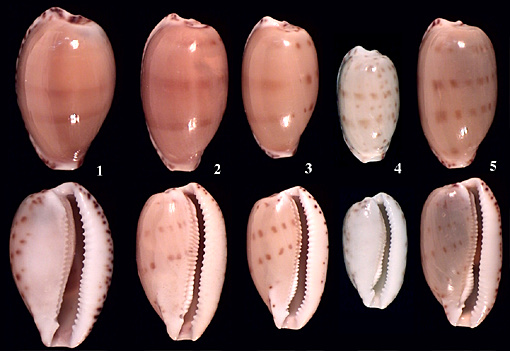
Little is known about the Notocypraea from deep water as they are small and easily drop through the mesh of fishing nets. Sometimes, they can be found in the stomach of fish. The plate below shows a form of comptonii which I called "ereginae" in G3. It is found in the stomach of the Queen Morwong (Nemadactylus valenciennesi) caught at 60 to 100 m off Esperance, SW Australia. These shells (fig 2-3) differ consistently from shallow water comptonii (fig.1: from the Esperance Jetty at 6 m)) by the slender shell with two very fine transverse bands and reduced terminal blotches, a narrower aperture, finer teeth and an enormous fossula and coarsely denticulate columellar peristome. These features are more typical for pulicaria (fig. 4: from Albany Harbour at 4 m), which is also found in the stomach of those fish, alongside the "ereginae" form of comptonii. The pulicaria from these depths are larger, more inflated and have a coarser, more projecting fossula than their shallow water relatives. Schilder (when describing hartsmithi in 1967) had already noted that a strong fossula is an archaic feature in the Cypraeidae. Recently, studies of the holotype of subcarnea revealed that this name has been misinterpreted. It is a valid species restricted to Tasmania.
1: comptonii, typical shallow water form (Esperance Jetty); 2-3: comptonii, deep water "ereginae" form, ex pisce off 60-100m Esperance;
4: pulicaria, typical shallow water, Albany Harbour; 5: pulicaria deep water variation, ex pisce off 60-100m Esperance;
First uploaded 2003, updated 2021
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}